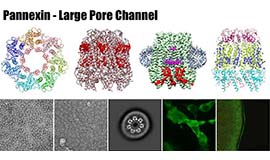
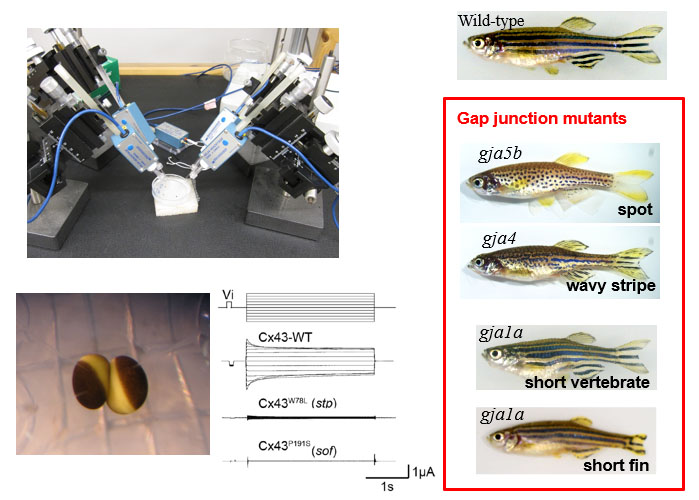
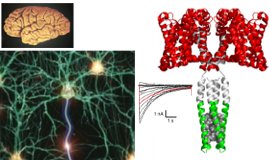
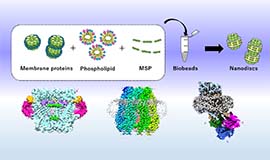
多細胞生物の多くは個体を形成する隣接細胞同士が結合構造で結ばれ、そこではシグナルのやり取りが行われています。この細胞間連絡を担う主要なコンポーネントの一つにギャップ結合という構造体があります。この実体は隣接する2つの細胞の細胞膜を貫通するギャップ結合チャネルが集積したもので、隣接細胞の細胞質を直接連絡することによって電気的、化学的なカップリングを実現しています。当研究室ではギャップ結合チャネルの生体内における多様な役割を構造研究に基づいて理解することを目指しています。クライオ電子顕微鏡を用いた高分解能構造解析と、機能解析や計算機科学を組み合わせて、ギャップ結合チャネルの開閉機構の解明に取り組んでいます。
関連ページ:大嶋